Citation Information
Silverman and Melamed, J Life Sci, Vol. 3, No. 3, March 2026:1-4 [https://doi.org/qtxq]
Introduction
Bacterial infections are becoming harder to treat. Antibiotics, which once seemed like miracle drugs, are slowly losing their power as bacteria evolve ways to resist them. This growing problem, called antibiotic resistance, is a major challenge in modern medicine. It is time to start investigating new ideas or maybe go back and take a look at our old tricks. One concept that is returning to the spotlight is using viruses that infect bacteria to overcome antibiotic-resistant bacteria. When we think about viruses, we usually think about getting sick. The flu virus, COVID-19, and many others infect our cells and make us feel miserable. But viruses don’t just infect humans. In fact, some viruses infect bacteria, and those viruses can be useful to us as they simply cannot infect the human cell due to their extremely specific nature. These bacteria-infecting viruses are called bacteriophages, or just phages, which literally means “bacteria eaters”. Phages are incredibly common
The hidden side of phage infection: RNA regulation
When a phage infects a bacterium, it follows a carefully planned sequence of steps. First, it must “land” on the bacterial surface: like a space module approaching the Moon and touching down at the precise landing site. The bacterial surface is not smooth or random; it has specific molecular “landing pads” that only certain phages can recognize. If the phage lands in the wrong place, the mission fails. Once the phage has successfully landed, it injects its genetic material into the bacterial cell, much like astronauts unloading their equipment after a successful touchdown. Inside the cell, the phage takes control of the bacterium’s machinery to build many copies of itself. Finally, the bacterial cell bursts open, releasing newly assembled phages that move on to infect neighboring bacterial cells.
For decades, scientists have studied how phages manipulate bacteria. Most of what we know involves proteins - molecular machines that phages use to shut down bacterial defences or redirect the cell’s resources. However, cells also use another powerful form of regulation using RNA molecules. Some RNA molecules are not used to make proteins at all. Instead, they act as regulators. These molecules, called small RNAs (sRNAs)
Could a phage also use small RNAs to manipulate its bacterial hosts?
For a long time, this question remained unanswered. Scientists lacked a global view of which bacterial and viral RNAs interact during infection. We reasoned that something was happening during viral invasion, but we needed to install a secret camera that could document all these interactions. Without this information, important regulatory networks could easily remain hidden.
To achieve this, we used a method called RIL-seq (RNA Interaction by Ligation and sequencing
listening in on RNA conversations
Using RIL-seq, we infected Escherichia coli (E. coli) with phage lambda, a classic model phage that has been studied for decades
What does this sRNA do?
As previously mentioned, sRNAs act by physically binding to other RNAs, resulting in their regulation. When taking a closer look at who PreS was interacting with on Hfq, we noticed that it binds to the RNA encoded by the bacterial gene dnaN. What really caught our attention was that dnaN only interacted with PreS. The dnaN gene encodes the β-sliding clamp
Here is the clever part: phage lambda does not bring its own DNA replication machinery. Instead, it relies on the bacterium’s system. That means anything that boosts bacterial DNA replication also helps the phage copy its own genome, and this is exactly what PreS does.
We found that PreS, while bound to Hfq, increases the production of the DnaN protein. It does so by helping ribosomes, the cell’s protein-making machines, start translating the dnaN mRNA more efficiently. This happens due to changes in the folding and structure of dnaN mRNA supporting our hypothesis.
In other words, PreS acts like an RNA switch that puts dnaN translation into high gear, just by binding to it. But we still needed to show that DNA replication is alleviated by PreS, so we synthetically overexpressed PreS from a plasmid in cells without any phage and showed that DNA synthesis was enhanced compared to an empty plasmid (Figure 1A). To neatly connect all the dots, we show that when infecting cells that have a mutant danN, fewer phages are produced and phage DNA quantity is lower. When PreS is present: The bacterium makes more sliding clamps, DNA replication becomes more efficient, phage DNA is copied faster, resulting in the production of more phage particles. Alternatively, when PreS is removed or mutated, everything slows down. Phage DNA replication is delayed (Figure 1B), and fewer viruses are made (Figure 1C).
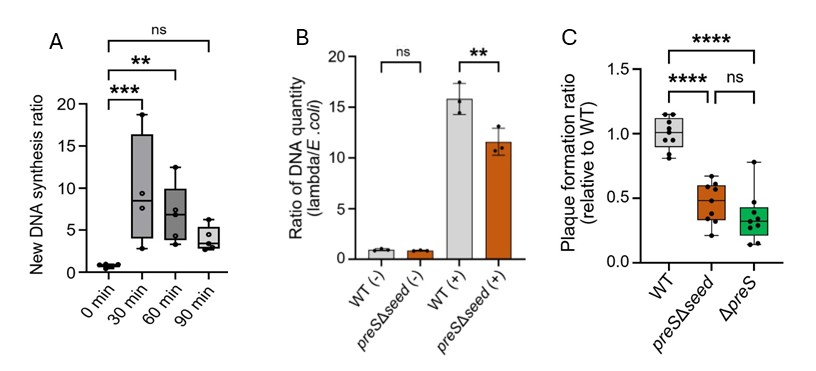
Figure 1 – PreS overexpression in E. coli results in enhanced DNA synthesis measured by BrdU incorporation into newly synthesized DNA (A). Results are relative to new DNA synthesis with an empty vector. Deletion of preS seed sequence resulted in reduced phage DNA quantity following induction of the lytic cycle by UV irradiation (B). Deletion of preS from lysogens results in reduced efficiency as measured by plaque assay of spontaneous lytic induction (C). All panels were adapted from Silverman A. et. al., 2025³
Why this discovery matters
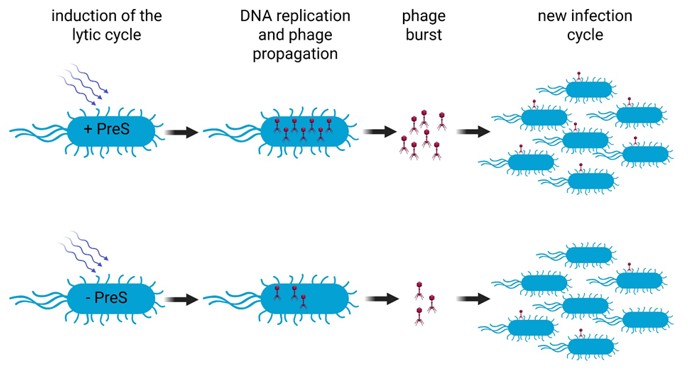
This work shows for the first time that a phage can directly control bacterial DNA replication using an sRNA. It reveals a completely new strategy that viruses use to hijack their hosts. Instead of smashing the system with brute force, the phage uses a subtle RNA-based trick, fine-tuning an essential bacterial process to serve its own needs (Figure 2). This changes how we think about phage biology. It suggests that many more phage-encoded sRNAs are waiting to be discovered, quietly pulling the strings inside infected cells.
By targeting dnaN, PreS pushes the bacterial replication system into high gear without completely disrupting the cell. This intermediate exploitation strategy allows the phage to extract maximum benefit from the host before the cell is destroyed. Understanding these strategies is also important for applied science. As phage therapy gains momentum, knowing how phages optimize their replication could help in selecting or engineering a phage that could work more efficiently against dangerous bacteria.
What's next?
There are still important pieces of the puzzle that we are trying to solve. First, we focused on one model bacterium (E. coli) and one well-studied phage (lambda). These are the “lab mice” of microbiology - they are extremely useful for discovery, but they do not represent all bacteria or all phage species found in nature. While we found that PreS-like sequences appear in related phages, we can only estimate how common this RNA-based strategy is across the enormous diversity of phages in the environment.
Second, although PreS interacts with many bacterial RNAs, and we have explored a few of its targets, there are many more RNA–RNA interactions. It is very likely that other sRNAs influence and contribute to phage biology, such as host-encoded sRNAs that could potentially create a counter-response to the infection. In other words, we have identified the control panel, but we have only tested a few of the buttons. There is still a lot to learn, and the complete control panel, although encrypted, is available in our research paper.
Acknowledgements
Work in the Melamed lab was supported by the ISF grants 826/22 and 2859/22. Aviezer Silverman is supported by the Abisch-Frenkel Foundation.
References
1. 1. Comeau AM, Hatfull GF, Krisch HM, Lindell D, Mann NH, Prangishvili D. Exploring the prokaryotic virosphere. Res Microbiol. 2008;159(5):306-313. doi:10.1016/j.resmic.2008.05.001
2. 2. Gordillo Altamirano FL, Barr JJ. Phage Therapy in the Postantibiotic Era. Clin Microbiol Rev. 2019;32(2). doi:10.1128/CMR.00066-18
3. 3. Silverman A, Nashef R, Wasserman R, et al. Phage-encoded small RNA hijacks host replication machinery to support the phage lytic cycle. Mol Cell. 2025;85(24):4678-4697.e12. doi:10.1016/j.molcel.2025.11.019
4. 4. Papenfort K, Melamed S. Small RNAs, Large Networks: Posttranscriptional Regulons in Gram-Negative Bacteria. Annu Rev Microbiol. 2023;77. doi:10.1146/annurev-micro-041320-025836
5. 5. Silverman A, Melamed S. Biological Insights from RNA–RNA Interactomes in Bacteria, as Revealed by RIL-seq. In: 2025:189-206. doi:10.1007/978-1-0716-4192-7_11
6. 6. Melamed S, Peer A, Faigenbaum-Romm R, et al. Global Mapping of Small RNA-Target Interactions in Bacteria. Mol Cell. 2016;63(5). doi:10.1016/j.molcel.2016.07.026
7. 7. Casjens SR, Hendrix RW. Bacteriophage lambda: Early pioneer and still relevant. Virology. 2015;479-480:310-330. doi:10.1016/j.virol.2015.02.010
8. 8. Stukenberg PT, Studwell-Vaughan PS, O’Donnell M. Mechanism of the sliding beta-clamp of DNA polymerase III holoenzyme. J Biol Chem. 1991;266(17):11328-11334.
Authors :

Dr. Melamed is a principal investigator at the Hebrew University of Jerusalem. He also serves at the director of the Microbiology track in the Faculty of Medicine at the Hebrew University of Jerusalem. Dr. Melamed is leading a group of researchers who are excited about RNA biology and love to do science. They aim to understand the roles played by regulatory RNAs in the relationships of bacteria with their environment.

Aviezer is a PhD student in the Melamed lab. His research work lead to their publication in Molecular Cell. The above layman summary is based on these published research findings. Please read more about the authors in the Mentor-Grad Student Spotlights, MGS, series.
Reviewers :

Dr. Aarzoo Grover is a postdoctoral associate at the University of Vermont in the Doublié lab studying the aspects of DNA damage and repair using biochemistry and structural biology. She is a postdoctoral reviewer at JoLS-Pub. Aarzoo coordinated the review of this summary

Cassandra is a Master of Pharmacology student at the University of Vermont. She also serves as a research technician in a structural biology laboratory, where she contributes to ongoing projects in molecular and biochemical characterization. Her research interests bridge pharmacology and neuroscience, and she intends to pursue a Ph.D. in Neuroscience following the completion of her master’s degree.