Citation Information
Schaefer and Bhosale, J Life Sci, Vol. 3, No. 2, June 2021:19-42 https://doi.org/gqnk
Mitochondria are increasingly recognized as key factors in cancer, affecting a multitude of tumor hallmarks. Likewise, sirtuin 3, a master regulator of mitochondrial function, has gained attention as a therapeutic target in cancer. As a main mediator of calorie restriction, sirtuin 3 inhibits anabolic reactions and cell proliferation, acting as a tumor suppressor. At the same time, sirtuin 3 protects the cell during nutrient stress by regulating cell death and DNA repair, thereby acting as an oncogene. Here we discuss this conflicting role of sirtuin 3 in cancer and evaluate the therapeutic potential of sirtuin 3 activators and inhibitors in different cancers.
Keywords: Sirtuin, SIRT3, Cancer, Mitochondria
Introduction
The lifetime risk of developing cancer is greater than 35% in the United States, and cancer is continuously among the three most common causes of death worldwide (1, 2). While tumors are very heterogeneous in their phenotype and underlying molecular changes, there are common characteristics that result in uncontrolled proliferation and malignancy. These hallmarks include proliferative signaling, genome instability, immune evasion, resistance to cell death and a deregulation of cellular energetics (3, 4).
The preference of tumors for anaerobic metabolism in the presence of oxygen, known as the Warburg effect, was discovered almost 100 years ago (5, 6). However, only within the last decade has a better understanding of this phenotype emerged. While oxidative phosphorylation is the most efficient with respect to ATP production, carbon is oxidized to CO2 and not available for biomass production. To maintain a high proliferation rate, tumor cells need to use glycolytic and TCA cycle intermediates such as Acetyl-CoA, amino acids, or ribose for biogenesis at the expense of efficient energy production (7, 8). These insights have resulted in a renewed focus on the role of mitochondria in cancer (9). Mitochondria are not only crucial in providing macromolecular precursors for cell proliferation but are also at the crossroads of many other hallmarks of cancer. Mitochondria are the major source of reactive oxygen species (ROS) in the cell, which can result in oxidative damage and mutations in DNA. Similarly, mitochondria regulate the antioxidative response (10) and DNA repair via the redox status and PARP1 (11), demonstrating a key role of mitochondria in genome stability. With respect to immune evasion, mitochondria were shown to regulate immune cell proliferation (12) and inflammatory response (13), suggesting a central role in the immune system and cancer-associated immune evasion (14, 15). Lastly, mitochondria are well known for their role in apoptotic cell death (16) and a dysregulation of mitochondrial apoptotic signaling was associated with cancer (17, 18). Taken together, mitochondria are a common link between the hallmarks of cancer. This emphasizes mitochondrial metabolism and its regulators, such as sirtuins, as promising targets for therapeutic intervention in cancer.
Sirtuins, or its homolog Sir-2, were first discovered in yeast and C. elegans in response to calorie restriction mediating an extended lifespan (19, 20). Sir-2 was described as a class III histone deacetylase, removing acetyl-groups from lysine residues on histones in an NAD+ dependent manner, thereby resulting in gene silencing (21).
In mammalian cells there are 7 sirtuin isoforms (SIRT1-SIRT7). They differ with respect to subcellular localization and substrate specificity. While SIRT1, 2, 6, 7 are predominantly localized in the nucleus or cytoplasm, SIRT3-5 are found mostly in the mitochondria (22). Sirtuins are not confined to histones as substrates but can perform posttranslational modifications on many non-histone proteins, resulting in either activation or inhibition of these enzymes. While deacetylation is the predominant modification mediated by sirtuins, some isoforms such as SIRT6 can remove larger Acyl-groups (23) or are involved in ADP-ribosylation (24).
All sirtuins display two core domains, a Rossmann fold domain common in nucleotide binding enzymes (25) and a zinc binding domain (26). Due to their NAD+ dependency, sirtuins function as cellular energy sensors detecting a low energy state by an increased NAD+/NADH ratio (19, 27), thereupon modifying gene expression or enzyme activity. Sirtuin activation generally results in a downstream induction of catabolism along with a reduction in anabolic reactions. In addition to their function as energy sensors and metabolic regulators (28), sirtuins were shown to be involved in cellular stress (29), the antioxidative response (22,30), inflammation (31,32) and DNA repair (33,34). Taken together, sirtuins are suggested to be a central hub for mitochondrial-nuclear interactions and for regulating cellular metabolism.
Dysregulation of the sirtuins has been associated with many disorders, such as aging (20,22,35), type II diabetes (36,37), cardiovascular disease (38-40), neurodegeneration (41,42), and cancer (43,44). With respect to cancer, it was demonstrated that several sirtuin knockout mice were more prone to develop tumors (45-47) suggesting sirtuin activation as a promising therapeutic target in cancer. However, recent evidence indicates a more conflicting role of sirtuins in tumorigenesis (48,49) with some tumors showing an upregulation of sirtuin expression (50,51). Given the crucial role of mitochondria in cancer, SIRT3 is of paramount interest since it is the predominant mitochondrial sirtuin. In this review we focus on SIRT3 aiming to unravel its dichotomous role in cancer, and evaluate activators and inhibitors of SIRT3 as therapeutic interventions in different malignancies.
SIRT3
SIRT3 is the major mitochondrial sirtuin and plays a key role in regulating mitochondrial enzymes via acetylation (52,53). It is found in most cell types and tissues with the highest levels in kidney, brain, heart, and liver (54). It is expressed as a 44kDa cytosolic protein containing an N-terminal mitochondrial targeting sequence. Proteolytic processing by the mitochondrial processing peptidase removes 101 amino acids on the N-terminus thereby resulting in activation of SIRT3 in the mitochondria. It is debated whether SIRT3 is exclusively found in the mitochondria (55) or if it translocates from the nucleus to the mitochondria upon cell stress (56). It appears clear that SIRT3 mainly plays a role in mitochondrial adaption to stress (57).
Regulation of SIRT3
Different stressors are known to regulate SIRT3 such as metabolic, oxidative, or genotoxic stress (58). Transcriptionally, an antioxidant response element upstream of SIRT3 was reported to regulate SIRT3 expression via the Nrf2-Keap axis in response to metabolic stress (59). On the protein level, it was suggested that SIRT3 can be targeted for degradation by the E3 ligase in response to DNA damage(60). In addition, PARP1 activation can deplete SIRT3 of NAD+ given a much lower Km of PARP1 for NAD+ (61,62). It is well known that NAD+ levels play an important role in stimulating SIRT3 activity (63). Conversely, nicotinamide, a product of the sirtuin reaction, was shown to mediate a feedback inhibition by binding to the C-pocket (64). There is some controversy with respect to sirtuin sensitivity for NADH and whether sirtuins sense the NAD+/nicotinamide ratio or the NAD+/NADH ratio (65). Interestingly, while sirtuin activity is stimulated by NAD+, sirtuin expression correlates with a low NAD+/NADH ratio (66). This suggests a differential effect of nuclear versus mitochondrial NAD+/NADH redox state on SIRT3 expression and activity respectively.
Physiological Function and Substrates of SIRT3
The main function of SIRT3 is the regulation of mitochondrial energy metabolism. SIRT3 deacetylates and thereby activates several enzymes of the TCA cycle such as pyruvate dehydrogenase (67), isocitrate dehydrogenase (IDH) (68) or Acetyl-CoA synthetase 2 (69). Additionally, it activates the mitochondrial electron transport system directly by deacetylating NDUFA9, a complex I subunit (70), and succinate dehydrogenase (71), complex II of the electron transport chain. Given the role of SIRT3 in metabolic adaptation to nutrient deprivation, SIRT3 stimulates fatty acid oxidation by activating long acyl chain dehydrogenases (57) and ketogenesis by activating the rate limiting enzyme 3-hydroxy-3-methylglutaryl-CoA synthase (72). Similarly, amino acid oxidation (73) and the associated urea cycle are activated by SIRT3 (74). At the same time, SIRT3 helps to preserve nutrient availability by limiting glucose uptake via Cyclophilin D (CYP-D) activation and subsequent dissociation of hexokinase 2 from the mitochondria (75). Taken together, SIRT3 stimulates catabolism and mediates a metabolic switch from glycolysis to oxidative phosphorylation.
Cell and nutrient stress as well as increased activity of the electron transport system are associated with increased ROS production. SIRT3 promotes the antioxidant response by activating Manganese Superoxide Dismutase (MnSOD) directly via deacetylation (76) and indirectly via FOXO3a-mediated upregulation (77). In addition, it stimulates NADPH production by IDH (68) and inactivates glutamic-oxaloacetic transaminase 2 (GOT2), thereby blocking the malate-aspartate shuttle (78). This maintains reducing equivalents in the cytosol, boosting the glutathione antioxidant system.
>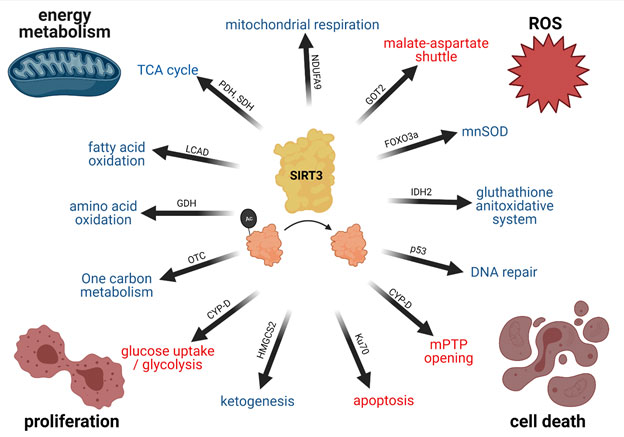
Figure 1: SIRT3 regulates Cell Metabolism and Health
SIRT3 deacetylates many mitochondrial proteins, thereby regulating a multitude of cellular functions. The functional cornerstones are an upregulation (indicated by blue) of mitochondrial energy metabolism, a downregulation (indicated by red) of reactive oxygen species, prevention of cell death and an inhibition of proliferation. Selected deacetylation targets mediating the physiological functions of SIRT3 are indicated on the arrows. Created with BioRender.com.
SIRT3 also plays a role in regulating DNA damage and cell death. SIRT3 was reported to deacetylate p53, which promotes its proteasomal degradation (79). Conversely, SIRT3 prevents degradation of 8-oxoguanine-DNA glycosylase 1 (80), a crucial enzyme in mitochondrial DNA repair. With respect to cell death, it was reported that SIRT3 activates Ku70, which subsequently binds Bax and prevents stress-induced cell death (81). Similarly, opening of the mitochondrial transition pore by CYP-D inactivation is reduced (82) preventing cell death due to mitochondrial calcium overload.
In summary, SIRT3 orchestrates the adaptation of mitochondrial energy metabolism. Upon sensing a low energy state via the redox ratio, it stimulates catabolic reactions and mitochondrial energy production. At the same time, it downregulates cell growth/replication but also protects the cell from ROS, DNA damage, or cell death (Figure 1).
SIRT3 in Cancer
Cancer cells can be distinguished from normal cells based on several key characteristics such as insensitivity to growth inhibitory signals and cell death resistance mechanisms, increased proliferative signaling, sustained angiogenesis, deregulation of metabolic pathways, genomic instability, increased inflammation, and tumor cell mobility (4). As such, deregulation of several physiological functions mediated by SIRT3 implicate a role for the mitochondrial sirtuin in cancer development. This association was underlined by an early study in SIRT3 knockout mice that demonstrated increased development of ER/PR-positive mammary tumors (45). SIRT3 deregulation has further been observed across several tumor subtypes. Mitochondrial SIRT3 can act as a tumor promoter or suppressor depending on the tumor profile and distinct molecular signatures of the cancer cells (Figure 2).
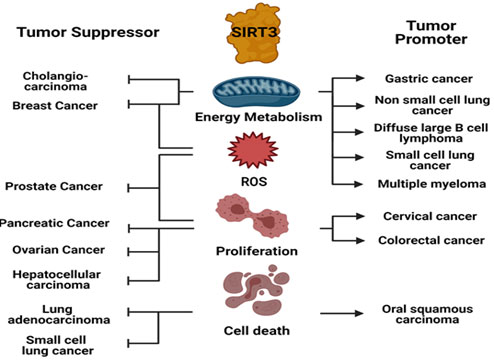
Figure 2: SIRT3 as Tumor Suppressor or Oncogene in different Tumors
SIRT3 mediates its tumor suppressor or oncogenic function by modulating cellular physiology differentially in a variety of tumors. Created with BioRender.com.
SIRT3 as Tumor Suppressor
A strong reliance on glycolytic energy metabolism to produce substrates for anabolic reactions is a hallmark of many tumors. SIRT3, in contrast, promotes a switch towards oxidative phosphorylation, thereby acting as a tumor suppressor. For example, in breast carcinoma SIRT3 inhibits glycolysis by deacetylating cyclophilin D, which leads to the dissociation of hexokinase II from the outer mitochondrial membrane protein voltage-dependent anion channel (83). Additionally, SIRT3-mediated stabilization of p53 via PTEN and MDM2 was shown to inhibit glycolysis and tumor cell proliferation by positive transcriptional regulation of genes involved in mitochondrial oxidative phosphorylation (84-86). A recent study in cholangiocarcinoma cells demonstrates the anti-Warburg effect of SIRT3 by inhibition of the hypoxia inducible factor 1α (HIF-1α)/pyruvate dehydrogenase kinase 1 (PDK1) pathway (87). SIRT3-mediated ROS depletion is also known to promote tumor inhibition. A study in SIRT3 knockout primary mouse embryonic fibroblasts demonstrated increased activation of HIF-1α, which is involved in transcriptional regulation of several glycolytic genes known to promote tumorigenesis (88,89). Activation of prolyl hydroxylases by SIRT3 was found to be responsible for destabilization of HIF-1α and mitigated its tumor promoting effects (90,91). Moreover, studies in prostate cancer reported that ROS depletion by SIRT3 could suppress the PI3K/Akt pathway resulting in ubiquitination and degradation of oncoprotein c-Myc, thus inhibiting tumor progression (92).
SIRT3 does not only influence proliferation via the abundance of glycolytic intermediates, but also regulates multiple pathways associated with proliferation, cell migration, and renewal. Given the role of SIRT3 in caloric restriction and downregulation of anabolic reactions a tumor suppressor role would be expected. SIRT3 mediated inactivation of glutamate oxaloacetate transaminase 2 in pancreatic cancer was shown to arrest proliferation and tumor growth (78). Deactivation of the proto-oncogene F-box protein S-phase kinase associated protein 2 by SIRT3 could also inhibit cancer cell progression and migration (93). SIRT3 was found to prevent hyperacetylation of enoyl-CoA hydratase 1 (ECHS1) in cancer, thereby inhibiting mTOR regulated proliferation (94). Furthermore, modulation of ROS signaling by SIRT3 was found to be relevant to tumor cell migration. Depletion of ROS levels by SIRT3 demonstrated diminished focal adhesion kinase (FAK) phosphorylation, which is required for breast cancer metastasis (95). SIRT3 was also shown to inhibit epithelial-to-mesenchymal transition (EMT) and metastasis in ovarian cancer cells by downregulation of Twist (96). Upregulated levels of SIRT3 in metastatic prostate cancer were shown to induce the inhibition of Wnt/β-catenin pathway, a central regulator of EMT and tumor metastasis (97). Additionally, SIRT3 mediated inhibition of cellular migration and metastasis have been demonstrated in hepatocellular carcinoma (98) and pancreatic ductal cell carcinoma (99).
Tumor-modulating effects of SIRT3 can also be attributed to its regulation of DNA repair and programmed cell death. As a tumor suppressor SIRT3 works to promote apoptosis. In HepG2 cells SIRT3 was found to induce Bax and Fas regulated apoptosis. This was mechanistically shown to be an effect of SIRT3-mediated upregulation of p53 and MnSOD (100). SIRT3 was also found to induce apoptosis in hepatocellular carcinoma by deacetylation of glycogen synthase kinase-3β, which further resulted in increased expression and mitochondrial translocation of Bax (101). Additionally, downregulation of Bcl-2 and JNK2 by SIRT3 was shown to contribute to apoptosis in hepatocellular carcinoma (102). In human lung adenocarcinoma tissue upregulated levels of SIRT3 results in increased translocation of apoptosis inducing factor (AIF) to the nucleus with higher levels of p53, p21, and depletion of ROS. This ultimately results in increased apoptosis (103). A recent study in small-cell lung cancer demonstrated that SIRT3 can inhibit tumor progression by promoting apoptosis and necroptosis via modulation of ubiquitination-mediated proteasomal degradation of mutant p53 protein (104). Ovarian cancer cell lines with increased expression of SIRT3 have also demonstrated increased sensitivity to metformin-induced apoptosis (105).
SIRT3 as Oncogene
Numerous studies over the past decade have also demonstrated the tumor promoting effects of SIRT3 across different cancers (Figure 2). This controversial role of SIRT3 as an oncogene likely arises from differential regulation of SIRT3-mediated cellular pathways in certain tumor settings. For example, increased protein and mRNA levels of SIRT3 were reported in non-small cell lung cancer tissues, and SIRT3 was found to regulate the activation of Akt promoting tumor malignancy (106). PTEN-deficient lung tissues have also demonstrated increased levels of SIRT3, which correlate with depleted p53 levels. SIRT3 was found to be responsible for ubiquitination and proteasomal degradation of p53 (79). SIRT3 was reported to activate lactate dehydrogenase in gastric cancer cells resulting in glycolysis and promotion of tumorigenesis (107). Hematological cancers that depend on oxidative phosphorylation also demonstrate an oncogenic role for SIRT3 (108). SIRT3 was found to promote lymphomagenesis in diffuse large B cell lymphomas by regulating TCA cycle via glutamate dehydrogenase (109). SIRT3 was further shown to deacetylate IDH2, a critical enzyme participating in forward Krebs cycle. The activation of IDH2 by SIRT3 promotes carcinogenesis in multiple myeloma cell lines (110).
SIRT3 can also participate in stimulation of proliferative signaling pathways and promote tumorigenesis. For example, SIRT3-mediated activation of nicotinamide mononucleotide adenylyl transferase 2 (NMNAT2) was shown to result in increased cellular proliferation and glycolytic flux (111). Similarly, SIRT3 activates the pyrroline-5-carboxylate reductase 1 (PYCR1) thereby stimulating proliferation (112). An increased expression of SIRT3 in lymph node positive breast cancer metastasis is also believed to contribute to the survival of aggressive cancer phenotypes. SIRT3-mediated deacetylation of serine hydroxymethyl-transferase 2 (SHMT2) was demonstrated to inhibit its lysosome-dependent degradation and contributed to colorectal cancer development (113). Other studies in colorectal cancer have demonstrated that SIRT3 activates the Akt/PTEN pathway, regulates transcription of metastatic genes like EGFR and BRAF, and promotes cancer cell migration and proliferation (114). Likewise, a study in cervical cancer cells revealed that SIRT3 deacetylated Acetyl-CoA carboxylase and reprogrammed fatty acid metabolism to promote migration and invasion (115).
As a central mitochondrial deacetylase, SIRT3 can respond to metabolic and genotoxic stress and protect cancer cells from apoptosis. SIRT3 was shown to deacetylate and bind with Ku70, which augmented Ku70-Bax interactions and prevented translocation of Bax to the mitochondria. This resulted in inhibition of apoptosis in HeLa cells (81). Overexpression of SIRT3 in oral squamous carcinoma cells is also shown to correlate with anoikis, apoptosis triggered by loss of extracellular matrix, and cell survival. Increased expression of SIRT3 in anoikis-resistant oral cancer was found to be accompanied by lower expression of receptor interacting protein, which can act as a negative regulator of SIRT3 (116). SIRT3-mediated deacetylation of histone H3 was also found to promote nonhomologous end joining repair. Histone H3 is known to be involved in the response to DNA damage and is found to be colocalized with γH2AX implicating a role for SIRT3 in maintenance of genomic stability in tumors (117). Likewise, another study reported that SIRT3 modulates the activity of human 8-oxoguanine-DNA glycosylase 1, a DNA repair enzyme, and assists in the response to mitochondrial DNA damage preventing stress mediated apoptosis (80).
Taken together, SIRT3 can act as a tumor suppressor or oncogene dependent on the specific tumor by regulating important tumor hallmarks such as energy metabolism, ROS, apoptosis, and proliferation.
Therapeutic targeting of SIRT3 in Cancer
As SIRT3 is implicated in the pathogenesis of cancer and other diseases, numerous compounds have been identified and studied for their activity on SIRT3. These molecules can regulate the activity of SIRT3 by either influencing its expression or by modulating its deacetylation function (118). Based on their effect, SIRT3 modulators can be broadly classified as SIRT3 activators or inhibitors (Table 1) (119).
SIRT3 Activators
Given the tumor suppressive function of SIRT3 across several cancer types, SIRT3 activation can prove to be an effective strategy. Endogenous regulators like nicotinamide riboside or nicotinamide mononucleotide can boost the intracellular levels of NAD+ and thereby stimulate SIRT3 (120). For example, it has been shown that nicotinamide mononucleotide can inhibit tumor growth and metastasis in mice (121). However, these endogenous regulators are not specific to SIRT3 but activate all sirtuins and directly affect cellular energy metabolism independent of the sirtuins.
Among the other compounds studied for specific SIRT3 activation, Honokiol, isolated from the bark of Magnolia officinalis, has shown great efficacy in the activation of SIRT3 (122). In vitro and in vivo studies with Honokiol in cancers of the breast, colon, prostate, liver, and squamous cell lung carcinomas have demonstrated promising results. However, most of its anti-cancer activity is attributed to its effects on other molecular targets such as NF-κb, STAT3, EGFR, mTOR, β-catenin and HIF1α, which are also known to be involved in pathways of cell survival, proliferation, and metabolism (123-128). Similarly, Silybin demonstrates weak SIRT3 activation (129,130) and has been found to suppress tumor growth (131,132), however a connection between both functions of Silybin have not yet been established. Resveratrol (119, 133, 134) and Piceatannol (133, 135,136) were found to activate most sirtuins but there are controversial reports of activation or inhibition of SIRT3 (137,138) and conclusive evidence supporting their role in SIRT3 mediated anti-cancer activity is lacking. More research into the discovery of synthetic, specific SIRT3 activators is needed to reveal a comprehensive overview of the potential of SIRT3 activation in different cancers as current activators lack specificity.
SIRT3 Inhibitors
On the contrary, improved understanding of the deacetylation process and identification of the crystal structure of SIRT3 has prompted research into the development of SIRT3 inhibitors. Using high throughput and in silico screening, several molecules with a wide range of core structures have been identified for SIRT3 inhibition. Of the numerous strategies employed, catalytic mechanism- based inhibition of SIRT3 has been widely studied. Nicotinamide serves as a natural sirtuin inhibitory molecule by binding to a conserved region in the sirtuin catalytic site and promoting a reverse base-exchange reaction instead of the deacetylation reaction (139). The chemo-preventive potential of nicotinamide was also demonstrated in keratinocyte cancers (140). However, the non-specific inhibition of all sirtuin isoforms and its rapid conversion to NAD+ limit the potential of nicotinamide for use in the treatment of cancer and other diseases. Thus, nicotinamide analogues have been developed for possible sirtuin inhibition. 3-TYP is a highly specific SIRT3 inhibitor (141), whereas Selisistat (or EX-527), a SIRT1, SIRT2 and SIRT3 inhibitor, has shown to work synergistically with Wee1 in lung cancer cells to promote growth inhibition and apoptosis (142-144).
Another strategy for the development of sirtuin inhibitors employs substrate specific competition. 4’-bromo resveratrol, an analogue of Resveratrol, was found to suppress SIRT1 and SIRT3 in melanoma where the targeted sirtuins had a proliferative role (145,146). It was observed that this dual inhibition of SIRT1 and SIRT3 was accompanied by induction of caspase-dependent apoptosis, inhibition of migratory potential, arrest in G1 phase, and ablation of aerobic glycolysis (146). Several SIRT3 inhibitors have been developed using a SIRT3-structure based approach but their biological activity has not been reported (147).
Chemical library screening-based development of SIRT3 inhibitors is another efficient strategy to identify novel SIRT3 inhibitors. A study utilizing self-assembled monolayer desorption/ionization mass spectrometry (SAMDI-MS) was used to screen small molecules for SIRT3 inhibition activity. This led to the discovery of SDX-437, which has an IC50 of 700nM and is one of the strongest SIRT3 inhibitors identified so far but has not yet been tested for potential anti-cancer activity (148). In contrast, Tenovin-6, a p53 activator and non-competitive inhibitor of SIRT3, has shown anti-tumor activity mostly through regulation of SIRT1, SIRT2 and to a lesser extent through SIRT3 (149,150). Minnelide, a water-soluble prodrug for diterpene tri-epoxide triptolide, was shown to inhibit the expression of SIRT3 in p53-deficient cancer cells and promoted mitochondrial dysfunction and upregulation of pro-apoptotic genes via mitigation of NF-κB signaling (151,152).
Table 1: Activators and Inhibitors of SIRT3
|
Compound |
Chemical Structure |
Mechanism/Specificity |
Applications |
|
SIRT3 Activators | |||
|
Honokiol |
|
Increased SIRT3 levels and activity (122) Not sirtuin-specific (123) |
Cardiomyopathy (122) Multiple cancers (123) |
|
Silybin |
|
Increased SIRT3 expression Not sirtuin-specific (129) |
Acute kidney injury (129) Liver disease (130) |
|
Nicotinamide Riboside |
|
Boosts NAD+ levels Activates all sirtuins alters energy metabolism |
Neurodegeneration Cancer Mitochondrial disorders (120) |
|
SIRT3 Inhibitors | |||
|
Nicotinamide |
|
Feedback inhibition (139) |
Multiple cancers (140) |
|
3-TYP |
|
NAD+ competitive SIRT3 inhibitor (141) |
Acute myeloid leukemia (156) |
|
Selisistat |
|
Occupies NAD+ binding pocket, Inhibitor of SIRT1, SIRT2, SIRT3 (143) |
Lung Cancer (142) Huntington’s disease (144) |
|
4-Bromo-Resveratrol |
|
Substrate competitive SIRT1 and SIRT3 inhibitor (145) |
Melanoma (146) |
|
SDX-437 |
|
Inhibitor of SIRT3 > SIRT1 |
- |
|
Tenovin-6 |
|
SIRT1, SIRT2 > SIRT3 inhibitor (149) |
Gastric cancer (150) |
|
Minnelide |
|
Regulates SIRT3 expression and proteasomal degradation (152) |
Non-small cell lung carcinoma (151) |
|
2-Methoxy-estradiol |
|
Binds to canonical and allosteric inhibitor binding sites of SIRT3 (153) |
Osteosarcoma (153) |
2-Methoxyestradiol, a natural derivative of 17β-estradiol was also found to inhibit the activity of SIRT3 by binding to both the canonical and allosteric inhibitor binding sites ultimately causing inhibition of mitochondrial biogenesis in osteosarcoma cancer cell model (153).
The use of Encoded library technology for screening of DNA encoded small molecules identified pan-inhibitors of SIRT1/2/3 such as carboxamides, which block the nicotinamide binding site (154). Additionally, the application of techniques such as DNA encoded Dynamic combinatorial library has led to the discovery of more selective and potent SIRT3 inhibitory ligands compared to the pan-sirtuin inhibitors (155). However, most of these compounds have yet to be evaluated for their efficacy in experimental models of cancer.
Taken together, the discovery of new and improved compounds targeting SIRT3 activity for therapeutic benefit is highly desired. Application of scientific drug design strategies will be particularly important to achieve this.
Perspective
SIRT3 is a crucial conductor of mitochondrial activity and cell metabolism thereby regulating important hallmarks of cancer. Alterations in SIRT3 and its downstream pathways have been found in many tumors emphasizing its involvement in cancer. However, SIRT3 cannot be pinpointed as a universal tumor suppressor or oncogene (48). One future challenge will be to elucidate the role of SIRT3 in different types and stages of tumors and to identify molecular tumor profiles that determine the divergent role of SIRT3. For example, SIRT3 generally acts more as a tumor suppressor in tumors that heavily rely on glycolysis as opposed to its oncogenic role in lymphomas, which rely more on oxidative energy metabolism. It could also be hypothesized that SIRT3 activation might work well preventing tumor growth in early stages due to its antiproliferative effect but is less effective in later stages because of its protective effect on cell stress. Future studies correlating the function of SIRT3 in specific tumors with their metabolic or expression profile will shed light on potential applications for SIRT3 modulators as cancer therapeutics in a personalized medicine approach (Figure 3).
Given the complex role of SIRT3 in cancer, application of SIRT3 activators or inhibitors needs to be carefully evaluated depending on the type and location of cancer. Application of SIRT3 activators might lead to increased carcinogenesis, whereas SIRT3 inhibition could culminate into multiple organ toxicities. As such, the development and application of targeted drug delivery systems might help overcome drawbacks associated with nonspecific effects of SIRT3 modulation. Similarly, combination of SIRT3 modulators with chemotherapeutics might be a promising approach given that SIRT3 can confer chemoresistance in some tumors (156). Despite the identification of several SIRT3 modulators, none have been approved for therapeutic use. Hence, an improved understanding of the contextual role of SIRT3 will be required for successful development and application of SIRT3 modulators as therapeutic anti-cancer drugs
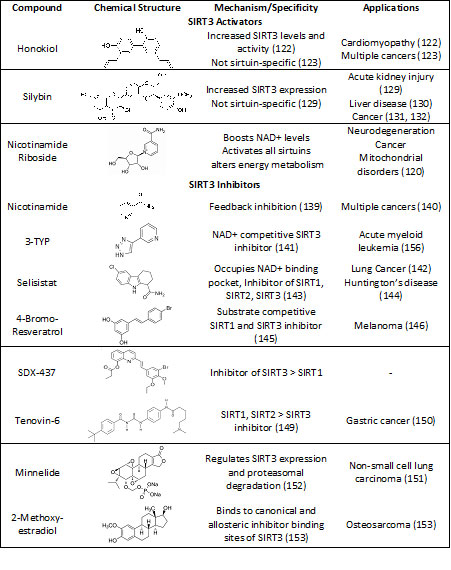
Figure 3: SIRT3 as potential therapeutic Target in Cancer
SIRT3 can act as either a tumor suppressor or oncogene. Combining the knowledge of molecular tumor signatures with a better understanding of SIRT3 biological function, regulation and the availability of SIRT3-specific compounds will allow evaluation of SIRT3 as therapeutic tumor target in future studies. Created with BioRender.com.
In addition to increasing our understanding of tumor modalities that favor SIRT3 modulation, more work is needed to understand the function and regulation of SIRT3. For example, there is still debate as to whether SIRT3 is regulated by NAD+ content, the NAD+/NA ratio or rather the NAD/NADH redox ratio (119). Given that SIRT3 is mostly located in the mitochondria, cytoplasmic and mitochondrial redox state might have differential effects on SIRT3. NADH imaging allows for assessing the redox state with subcellular resolution (157,158) and future studies combining this powerful technology with SIRT3 expression and activity promise crucial insights into SIRT3 regulation. This is especially important given the current limitation on SIRT3 activators. Most evidence is related to usage of nonspecific drugs, such as NAD+ boosting compounds, whose effects are only partially mediated through SIRT3. A better understanding of SIRT3 regulation could open new approaches for its activation. In addition, development of more specific, synthetic activators and inhibitors is essential to allow for targeted SIRT3 modulation in different tumor settings.
Taken together, SIRT3 demonstrates a high functional relevance in tumor biology but a better understanding of suitable tumor profiles and more specific compounds are necessary to fully evaluate the potential of SIRT3 modulation as a cancer therapeutic.
Summary
- Sirtuins are deacylases involved in epigenetic and posttranslational regulation
- Sirtuin 3 is a key regulator of mitochondrial function
- Sirtuin 3 is involved in tumor pathogenesis by regulating energy metabolism, ROS levels, proliferation and apoptosis
- Sirtuin 3 can act as tumor suppressor or oncogene dependent on the tumor
- Sirtuin 3 is a promising therapeutic target in personalized tumor therapy upon development of more specific activators and inhibitors.
Disclosures
No conflicts of interest, financial or otherwise, are declared by the authors.
Acknowledgements
The authors thank Dr. Douglas C. Wallace for his supervision and support in writing this review. We further thank Dr. Ryan Morrow and Arrienne Butic for proofreading the manuscript and providing constructive feedback.
Funding
This work was supported by the German Research Foundation (SCHA 2182/1-1).
References
|
1. Mattiuzzi C, Lippi G. Current Cancer Epidemiology. J Epidemiol Glob Health. 2019;9(4):217-22. https://doi.org/10.2991/jegh.k.191008.001 PMid:31854162 PMCid:PMC7310786 |
|
|
2. Henley SJ, Ward EM, Scott S, Ma J, Anderson RN, Firth AU, et al. Annual report to the nation on the status of cancer, part I: National cancer statistics. Cancer. 2020;126(10):2225-49. https://doi.org/10.1002/cncr.32802 PMid:32162336 PMCid:PMC7299151 |
|
|
3. Hanahan D, Weinberg RA. The hallmarks of cancer. Cell. 2000;100(1):57-70. https://doi.org/10.1016/S0092-8674(00)81683-9 |
|
|
4. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646-74. https://doi.org/10.1016/j.cell.2011.02.013 PMid:21376230 |
|
|
5. DeBerardinis RJ, Chandel NS. We need to talk about the Warburg effect. Nat Metab. 2020;2(2):127-9. https://doi.org/10.1038/s42255-020-0172-2 |
|
|
6. Warburg O, Wind F, Negelein E. THE METABOLISM OF TUMORS IN THE BODY. J Gen Physiol. 1927;8(6):519-30. https://doi.org/10.1085/jgp.8.6.519 |
|
|
7. Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science. 2009;324(5930):1029-33. https://doi.org/10.1126/science.1160809 PMid:19460998 PMCid:PMC2849637 |
|
|
8. Pavlova NN, Thompson CB. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016;23(1):27-47. https://doi.org/10.1016/j.cmet.2015.12.006 |
|
|
9. Wallace DC. Mitochondria and cancer. Nat Rev Cancer. 2012;12(10):685-98. https://doi.org/10.1038/nrc3365 PMid:23001348 PMCid:PMC4371788 |
|
|
10. Sies H, Berndt C, Jones DP. Oxidative Stress. Annu Rev Biochem. 2017;86:715-48. https://doi.org/10.1146/annurev-biochem-061516-045037 |
|
|
11. Herrmann GK, Russell WK, Garg NJ, Yin YW. Poly(ADP-Ribose) polymerase 1 regulates mitochondrial DNA repair in a metabolism-dependent manner. J Biol Chem. 2021:100309. https://doi.org/10.1016/j.jbc.2021.100309 PMid:33482196 PMCid:PMC7949115 |
|
|
12. Quinn WJ, Jiao J, TeSlaa T, Stadanlick J, Wang Z, Wang L, et al. Lactate Limits T Cell Proliferation via the NAD(H) Redox State. Cell Rep. 2020;33(11):108500. https://doi.org/10.1016/j.celrep.2020.108500 PMid:33326785 PMCid:PMC7830708 |
|
|
13. Green DR, Galluzzi L, Kroemer G. Mitochondria and the autophagy-inflammation-cell death axis in organismal aging. Science. 2011;333(6046):1109-12. |
|
|
14. Breda CNS, Davanzo GG, Basso PJ, Saraiva Câmara NO, Moraes-Vieira PMM. Mitochondria as central hub of the immune system. Redox Biol. 2019;26:101255. https://doi.org/10.1016/j.redox.2019.101255 PMid:31247505 PMCid:PMC6598836 |
|
|
15. Klein K, He K, Younes AI, Barsoumian HB, Chen D, Ozgen T, et al. Role of Mitochondria in Cancer Immune Evasion and Potential Therapeutic Approaches. Front Immunol. 2020;11:573326. https://doi.org/10.3389/fimmu.2020.573326 PMid:33178201 PMCid:PMC7596324 |
|
|
16. Bock FJ, Tait SWG. Mitochondria as multifaceted regulators of cell death. Nat Rev Mol Cell Biol. 2020;21(2):85-100. https://doi.org/10.1038/s41580-019-0173-8 PMid:31636403 |
|
|
17. Matsuura K, Canfield K, Feng W, Kurokawa M. Metabolic Regulation of Apoptosis in Cancer. Int Rev Cell Mol Biol. 2016;327:43-87. |
|
|
18. Plati J, Bucur O, Khosravi-Far R. Dysregulation of apoptotic signaling in cancer: molecular mechanisms and therapeutic opportunities. J Cell Biochem. 2008;104(4):1124-49. https://doi.org/10.1002/jcb.21707 PMid:18459149 PMCid:PMC2941905 |
|
|
19. Lin SJ, Defossez PA, Guarente L. Requirement of NAD and SIR2 for life-span extension by calorie restriction in Saccharomyces cerevisiae. Science. 2000;289(5487):2126-8. https://doi.org/10.1126/science.289.5487.2126 PMid:11000115 |
|
|
20. Tissenbaum HA, Guarente L. Increased dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans. Nature. 2001;410(6825):227-30. |
|
|
21. Imai S, Armstrong CM, Kaeberlein M, Guarente L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature. 2000;403(6771):795-800. https://doi.org/10.1038/35001622 PMid:10693811 |
|
|
22. van de Ven RAH, Santos D, Haigis MC. Mitochondrial Sirtuins and Molecular Mechanisms of Aging. Trends Mol Med. 2017;23(4):320-31. |
|
|
23. Chang AR, Ferrer CM, Mostoslavsky R. SIRT6, a Mammalian Deacylase with Multitasking Abilities. Physiol Rev. 2020;100(1):145-69. |
|
|
24. Aravind L, Zhang D, de Souza RF, Anand S, Iyer LM. The natural history of ADP-ribosyltransferases and the ADP-ribosylation system. Curr Top Microbiol Immunol. 2015;384:3-32. https://doi.org/10.1007/82_2014_414 PMid:25027823 PMCid:PMC6126934 |
|
|
25. Rossmann MG, Moras D, Olsen KW. Chemical and biological evolution of nucleotide-binding protein. Nature. 1974;250(463):194-9. |
|
|
26. Sanders BD, Jackson B, Marmorstein R. Structural basis for sirtuin function: what we know and what we don't. Biochim Biophys Acta. 2010;1804(8):1604-16. |
|
|
27. Schwer B, North BJ, Frye RA, Ott M, Verdin E. The human silent information regulator (Sir)2 homologue hSIRT3 is a mitochondrial nicotinamide adenine dinucleotide-dependent deacetylase. J Cell Biol. 2002;158(4):647-57. https://doi.org/10.1083/jcb.200205057 PMid:12186850 PMCid:PMC2174009 |
|
|
28. Chang HC, Guarente L. SIRT1 and other sirtuins in metabolism. Trends Endocrinol Metab. 2014;25(3):138-45. https://doi.org/10.1016/j.tem.2013.12.001 |
|
|
29. Brunet A, Sweeney LB, Sturgill JF, Chua KF, Greer PL, Lin Y, et al. Stress-dependent regulation of FOXO transcription factors by the SIRT1 deacetylase. Science. 2004;303(5666):2011-5. https://doi.org/10.1126/science.1094637 PMid:14976264 |
|
|
30. Kobayashi Y, Furukawa-Hibi Y, Chen C, Horio Y, Isobe K, Ikeda K, et al. SIRT1 is critical regulator of FOXO-mediated transcription in response to oxidative stress. Int J Mol Med. 2005;16(2):237-43. https://doi.org/10.3892/ijmm.16.2.237 PMid:16012755 |
|
|
31. Vachharajani VT, Liu T, Brown CM, Wang X, Buechler NL, Wells JD, et al. SIRT1 inhibition during the hypoinflammatory phenotype of sepsis enhances immunity and improves outcome. J Leukoc Biol. 2014;96(5):785-96. https://doi.org/10.1189/jlb.3MA0114-034RR PMid:25001863 PMCid:PMC4197566 |
|
|
32. Vachharajani VT, Liu T, Wang X, Hoth JJ, Yoza BK, McCall CE. Sirtuins Link Inflammation and Metabolism. J Immunol Res. 2016;2016:8167273. |
|
|
33. Mostoslavsky R, Chua KF, Lombard DB, Pang WW, Fischer MR, Gellon L, et al. Genomic instability and aging-like phenotype in the absence of mammalian SIRT6. Cell. 2006;124(2):315-29. https://doi.org/10.1016/j.cell.2005.11.044 PMid:16439206 |
|
|
34. Lagunas-Rangel FA. Current role of mammalian sirtuins in DNA repair. DNA Repair (Amst). 2019;80:85-92. https://doi.org/10.1016/j.dnarep.2019.06.009 |
|
|
35. Kanfi Y, Naiman S, Amir G, Peshti V, Zinman G, Nahum L, et al. The sirtuin SIRT6 regulates lifespan in male mice. Nature. 2012;483(7388):218-21. |
|
|
36. Banks AS, Kon N, Knight C, Matsumoto M, Gutiérrez-Juárez R, Rossetti L, et al. SirT1 gain of function increases energy efficiency and prevents diabetes in mice. Cell Metab. 2008;8(4):333-41. https://doi.org/10.1016/j.cmet.2008.08.014 PMid:18840364 PMCid:PMC3222897 |
|
|
37. Milne JC, Lambert PD, Schenk S, Carney DP, Smith JJ, Gagne DJ, et al. Small molecule activators of SIRT1 as therapeutics for the treatment of type 2 diabetes. Nature. 2007;450(7170):712-6. https://doi.org/10.1038/nature06261 PMid:18046409 PMCid:PMC2753457 |
|
|
38. Matsushima S, Sadoshima J. The role of sirtuins in cardiac disease. Am J Physiol Heart Circ Physiol. 2015;309(9):H1375-89. |
|
|
39. Oka S, Alcendor R, Zhai P, Park JY, Shao D, Cho J, et al. PPARα-Sirt1 complex mediates cardiac hypertrophy and failure through suppression of the ERR transcriptional pathway. Cell Metab. 2011;14(5):598-611. https://doi.org/10.1016/j.cmet.2011.10.001 PMid:22055503 PMCid:PMC3217210 |
|
|
40. Winnik S, Auwerx J, Sinclair DA, Matter CM. Protective effects of sirtuins in cardiovascular diseases: from bench to bedside. Eur Heart J. 2015;36(48):3404-12. https://doi.org/10.1093/eurheartj/ehv290 PMid:26112889 PMCid:PMC4685177 |
|
|
41. Buck E, Bayer H, Lindenberg KS, Hanselmann J, Pasquarelli N, Ludolph AC, et al. Comparison of Sirtuin 3 Levels in ALS and Huntington's Disease-Differential Effects in Human Tissue Samples vs. Transgenic Mouse Models. Front Mol Neurosci. 2017;10:156. https://doi.org/10.3389/fnmol.2017.00156 |
|
|
42. Lautrup S, Sinclair DA, Mattson MP, Fang EF. NAD. Cell Metab. 2019;30(4):630-55. https://doi.org/10.1016/j.cmet.2019.09.001 PMid:31577933 PMCid:PMC6787556 |
|
|
43. Chalkiadaki A, Guarente L. The multifaceted functions of sirtuins in cancer. Nat Rev Cancer. 2015;15(10):608-24. https://doi.org/10.1038/nrc3985 |
|
|
44. Roth M, Chen WY. Sorting out functions of sirtuins in cancer. Oncogene. 2014;33(13):1609-20. https://doi.org/10.1038/onc.2013.120 PMid:23604120 PMCid:PMC4295628 |
|
|
45. Kim HS, Patel K, Muldoon-Jacobs K, Bisht KS, Aykin-Burns N, Pennington JD, et al. SIRT3 is a mitochondria-localized tumor suppressor required for maintenance of mitochondrial integrity and metabolism during stress. Cancer Cell. 2010;17(1):41-52. https://doi.org/10.1016/j.ccr.2009.11.023 |
|
|
46. Kim HS, Vassilopoulos A, Wang RH, Lahusen T, Xiao Z, Xu X, et al. SIRT2 maintains genome integrity and suppresses tumorigenesis through regulating APC/C activity. Cancer Cell. 2011;20(4):487-99. https://doi.org/10.1016/j.ccr.2011.09.004 PMid:22014574 PMCid:PMC3199577 |
|
|
47. Sebastián C, Zwaans BM, Silberman DM, Gymrek M, Goren A, Zhong L, et al. The histone deacetylase SIRT6 is a tumor suppressor that controls cancer metabolism. Cell. 2012;151(6):1185-99. https://doi.org/10.1016/j.cell.2012.10.047 PMid:23217706 PMCid:PMC3526953 |
|
|
48. Boily G, He XH, Pearce B, Jardine K, McBurney MW. SirT1-null mice develop tumors at normal rates but are poorly protected by resveratrol. Oncogene. 2009;28(32):2882-93. https://doi.org/10.1038/onc.2009.147 PMid:19503100 |
|
|
49. Xiong Y, Wang M, Zhao J, Han Y, Jia L. Sirtuin 3: A Janus face in cancer (Review). Int J Oncol. 2016;49(6):2227-35. https://doi.org/10.3892/ijo.2016.3767 |
|
|
50. Jung-Hynes B, Nihal M, Zhong W, Ahmad N. Role of sirtuin histone deacetylase SIRT1 in prostate cancer. A target for prostate cancer management via its inhibition? J Biol Chem. 2009;284(6):3823-32. https://doi.org/10.1074/jbc.M807869200 PMid:19075016 PMCid:PMC2635052 |
|
|
51. Alhazzazi TY, Kamarajan P, Joo N, Huang JY, Verdin E, D'Silva NJ, et al. Sirtuin-3 (SIRT3), a novel potential therapeutic target for oral cancer. Cancer. 2011;117(8):1670-8. https://doi.org/10.1002/cncr.25676 PMid:21472714 PMCid:PMC3117020 |
|
|
52. Lombard DB, Alt FW, Cheng HL, Bunkenborg J, Streeper RS, Mostoslavsky R, et al. Mammalian Sir2 homolog SIRT3 regulates global mitochondrial lysine acetylation. Mol Cell Biol. 2007;27(24):8807-14. https://doi.org/10.1128/MCB.01636-07 PMid:17923681 PMCid:PMC2169418 |
|
|
53. Hebert AS, Dittenhafer-Reed KE, Yu W, Bailey DJ, Selen ES, Boersma MD, et al. Calorie restriction and SIRT3 trigger global reprogramming of the mitochondrial protein acetylome. Mol Cell. 2013;49(1):186-99. https://doi.org/10.1016/j.molcel.2012.10.024 PMid:23201123 PMCid:PMC3704155 |
|
|
54. Jin L, Galonek H, Israelian K, Choy W, Morrison M, Xia Y, et al. Biochemical characterization, localization, and tissue distribution of the longer form of mouse SIRT3. Protein Sci. 2009;18(3):514-25. https://doi.org/10.1002/pro.50 PMid:19241369 PMCid:PMC2760358 |
|
|
55. Cooper HM, Spelbrink JN. The human SIRT3 protein deacetylase is exclusively mitochondrial. Biochem J. 2008;411(2):279-85. |
|
|
56. Scher MB, Vaquero A, Reinberg D. SirT3 is a nuclear NAD+-dependent histone deacetylase that translocates to the mitochondria upon cellular stress. Genes Dev. 2007;21(8):920-8. https://doi.org/10.1101/gad.1527307 PMid:17437997 PMCid:PMC1847710 |
|
|
57. Hirschey MD, Shimazu T, Goetzman E, Jing E, Schwer B, Lombard DB, et al. SIRT3 regulates mitochondrial fatty-acid oxidation by reversible enzyme deacetylation. Nature. 2010;464(7285):121-5. https://doi.org/10.1038/nature08778 PMid:20203611 PMCid:PMC2841477 |
|
|
58. Marcus JM, Andrabi SA. SIRT3 Regulation Under Cellular Stress: Making Sense of the Ups and Downs. Front Neurosci. 2018;12:799. |
|
|
59. Satterstrom FK, Swindell WR, Laurent G, Vyas S, Bulyk ML, Haigis MC. Nuclear respiratory factor 2 induces SIRT3 expression. Aging Cell. 2015;14(5):818-25. https://doi.org/10.1111/acel.12360 PMid:26109058 PMCid:PMC4568969 |
|
|
60. Yoon SP, Kim J. Poly(ADP-ribose) polymerase 1 contributes to oxidative stress through downregulation of sirtuin 3 during cisplatin nephrotoxicity. Anat Cell Biol. 2016;49(3):165-76. https://doi.org/10.5115/acb.2016.49.3.165 PMid:27722009 PMCid:PMC5052225 |
|
|
61. Hirschey MD, Shimazu T, Jing E, Grueter CA, Collins AM, Aouizerat B, et al. SIRT3 deficiency and mitochondrial protein hyperacetylation accelerate the development of the metabolic syndrome. Mol Cell. 2011;44(2):177-90. https://doi.org/10.1016/j.molcel.2011.07.019 PMid:21856199 PMCid:PMC3563434 |
|
|
62. Cantó C, Sauve AA, Bai P. Crosstalk between poly(ADP-ribose) polymerase and sirtuin enzymes. Mol Aspects Med. 2013;34(6):1168-201. |
|
|
63. Onyango P, Celic I, McCaffery JM, Boeke JD, Feinberg AP. SIRT3, a human SIR2 homologue, is an NAD-dependent deacetylase localized to mitochondria. Proc Natl Acad Sci U S A. 2002;99(21):13653-8. https://doi.org/10.1073/pnas.222538099 PMid:12374852 PMCid:PMC129731 |
|
|
64. Bitterman KJ, Anderson RM, Cohen HY, Latorre-Esteves M, Sinclair DA. Inhibition of silencing and accelerated aging by nicotinamide, a putative negative regulator of yeast sir2 and human SIRT1. J Biol Chem. 2002;277(47):45099-107. https://doi.org/10.1074/jbc.M205670200 PMid:12297502 |
|
|
65. Madsen AS, Andersen C, Daoud M, Anderson KA, Laursen JS, Chakladar S, et al. Investigating the Sensitivity of NAD+-dependent Sirtuin Deacylation Activities to NADH. J Biol Chem. 2016;291(13):7128-41. https://doi.org/10.1074/jbc.M115.668699 PMid:26861872 PMCid:PMC4807294 |
|
|
66. Gambini J, Gomez-Cabrera MC, Borras C, Valles SL, Lopez-Grueso R, Martinez-Bello VE, et al. Free [NADH]/[NAD(+)] regulates sirtuin expression. Arch Biochem Biophys. 2011;512(1):24-9. https://doi.org/10.1016/j.abb.2011.04.020 PMid:21575591 |
|
|
67. Shan C, Kang HB, Elf S, Xie J, Gu TL, Aguiar M, et al. Tyr-94 phosphorylation inhibits pyruvate dehydrogenase phosphatase 1 and promotes tumor growth. J Biol Chem. 2014;289(31):21413-22. https://doi.org/10.1074/jbc.M114.581124 PMid:24962578 PMCid:PMC4118105 |
|
|
68. Someya S, Yu W, Hallows WC, Xu J, Vann JM, Leeuwenburgh C, et al. Sirt3 mediates reduction of oxidative damage and prevention of age-related hearing loss under caloric restriction. Cell. 2010;143(5):802-12. https://doi.org/10.1016/j.cell.2010.10.002 PMid:21094524 PMCid:PMC3018849 |
|
|
69. Hallows WC, Lee S, Denu JM. Sirtuins deacetylate and activate mammalian acetyl-CoA synthetases. Proc Natl Acad Sci U S A. 2006;103(27):10230-5. |
|
|
70. Ahn BH, Kim HS, Song S, Lee IH, Liu J, Vassilopoulos A, et al. A role for the mitochondrial deacetylase Sirt3 in regulating energy homeostasis. Proc Natl Acad Sci U S A. 2008;105(38):14447-52. https://doi.org/10.1073/pnas.0803790105 PMid:18794531 PMCid:PMC2567183 |
|
|
71. Cimen H, Han MJ, Yang Y, Tong Q, Koc H, Koc EC. Regulation of succinate dehydrogenase activity by SIRT3 in mammalian mitochondria. Biochemistry. 2010;49(2):304-11. https://doi.org/10.1021/bi901627u PMid:20000467 PMCid:PMC2826167 |
|
|
72. Shimazu T, Hirschey MD, Hua L, Dittenhafer-Reed KE, Schwer B, Lombard DB, et al. SIRT3 deacetylates mitochondrial 3-hydroxy-3-methylglutaryl CoA synthase 2 and regulates ketone body production. Cell Metab. 2010;12(6):654-61. https://doi.org/10.1016/j.cmet.2010.11.003 PMid:21109197 PMCid:PMC3310379 |
|
|
73. Schlicker C, Gertz M, Papatheodorou P, Kachholz B, Becker CF, Steegborn C. Substrates and regulation mechanisms for the human mitochondrial sirtuins Sirt3 and Sirt5. J Mol Biol. 2008;382(3):790-801. https://doi.org/10.1016/j.jmb.2008.07.048 PMid:18680753 |
|
|
74. Hallows WC, Yu W, Smith BC, Devries MK, Devires MK, Ellinger JJ, et al. Sirt3 promotes the urea cycle and fatty acid oxidation during dietary restriction. Mol Cell. 2011;41(2):139-49. https://doi.org/10.1016/j.molcel.2011.01.002 PMid:21255725 PMCid:PMC3101115 |
|
|
75. Wei L, Zhou Y, Dai Q, Qiao C, Zhao L, Hui H, et al. Oroxylin A induces dissociation of hexokinase II from the mitochondria and inhibits glycolysis by SIRT3-mediated deacetylation of cyclophilin D in breast carcinoma. Cell Death Dis. 2013;4:e601. https://doi.org/10.1038/cddis.2013.131 PMid:23598413 PMCid:PMC3641353 |
|
|
76. Tao R, Coleman MC, Pennington JD, Ozden O, Park SH, Jiang H, et al. Sirt3-mediated deacetylation of evolutionarily conserved lysine 122 regulates MnSOD activity in response to stress. Mol Cell. 2010;40(6):893-904. https://doi.org/10.1016/j.molcel.2010.12.013 PMid:21172655 PMCid:PMC3266626 |
|
|
77. Sundaresan NR, Gupta M, Kim G, Rajamohan SB, Isbatan A, Gupta MP. Sirt3 blocks the cardiac hypertrophic response by augmenting Foxo3a-dependent antioxidant defense mechanisms in mice. J Clin Invest. 2009;119(9):2758-71. https://doi.org/10.1172/JCI39162 PMid:19652361 PMCid:PMC2735933 |
|
|
78. Yang H, Zhou L, Shi Q, Zhao Y, Lin H, Zhang M, et al. SIRT3-dependent GOT2 acetylation status affects the malate-aspartate NADH shuttle activity and pancreatic tumor growth. EMBO J. 2015;34(8):1110-25. https://doi.org/10.15252/embj.201591041 PMid:25755250 PMCid:PMC4406655 |
|
|
79. Xiong Y, Wang L, Wang S, Wang M, Zhao J, Zhang Z, et al. SIRT3 deacetylates and promotes degradation of P53 in PTEN-defective non-small cell lung cancer. J Cancer Res Clin Oncol. 2018;144(2):189-98. https://doi.org/10.1007/s00432-017-2537-9 PMid:29103158 |
|
|
80. Cheng Y, Ren X, Gowda AS, Shan Y, Zhang L, Yuan YS, et al. Interaction of Sirt3 with OGG1 contributes to repair of mitochondrial DNA and protects from apoptotic cell death under oxidative stress. Cell Death Dis. 2013;4:e731. https://doi.org/10.1038/cddis.2013.254 PMid:23868064 PMCid:PMC3730425 |
|
|
81. Sundaresan NR, Samant SA, Pillai VB, Rajamohan SB, Gupta MP. SIRT3 is a stress-responsive deacetylase in cardiomyocytes that protects cells from stress-mediated cell death by deacetylation of Ku70. Mol Cell Biol. 2008;28(20):6384-401. https://doi.org/10.1128/MCB.00426-08 PMid:18710944 PMCid:PMC2577434 |
|
|
82. Hafner AV, Dai J, Gomes AP, Xiao CY, Palmeira CM, Rosenzweig A, et al. Regulation of the mPTP by SIRT3-mediated deacetylation of CypD at lysine 166 suppresses age-related cardiac hypertrophy. Aging (Albany NY). 2010;2(12):914-23. https://doi.org/10.18632/aging.100252 PMid:21212461 PMCid:PMC3034180 |
|
|
83. Pedersen PL. Warburg, me and Hexokinase 2: Multiple discoveries of key molecular events underlying one of cancers' most common phenotypes, the "Warburg Effect", i.e., elevated glycolysis in the presence of oxygen. J Bioenerg Biomembr. 2007;39(3):211-22. https://doi.org/10.1007/s10863-007-9094-x PMid:17879147 |
|
|
84. Dalmonte ME, Forte E, Genova ML, Giuffrè A, Sarti P, Lenaz G. Control of respiration by cytochrome c oxidase in intact cells: role of the membrane potential. J Biol Chem. 2009;284(47):32331-5. https://doi.org/10.1074/jbc.M109.050146 PMid:19776013 PMCid:PMC2781647 |
|
|
85. Zhao W, Wang J, Varghese M, Ho L, Mazzola P, Haroutunian V, et al. Impaired mitochondrial energy metabolism as a novel risk factor for selective onset and progression of dementia in oldest-old subjects. Neuropsychiatr Dis Treat. 2015;11:565-74. https://doi.org/10.2147/NDT.S74898 PMid:25784811 PMCid:PMC4356684 |
|
|
86. Shen L, Sun X, Fu Z, Yang G, Li J, Yao L. The fundamental role of the p53 pathway in tumor metabolism and its implication in tumor therapy. Clin Cancer Res. 2012;18(6):1561-7. https://doi.org/10.1158/1078-0432.CCR-11-3040 PMid:22307140 |
|
|
87. Xu L, Li Y, Zhou L, Dorfman RG, Liu L, Cai R, et al. SIRT3 elicited an anti-Warburg effect through HIF1α/PDK1/PDHA1 to inhibit cholangiocarcinoma tumorigenesis. Cancer Med. 2019;8(5):2380-91. https://doi.org/10.1002/cam4.2089 PMid:30993888 PMCid:PMC6536927 |
|
|
88. Bell EL, Emerling BM, Ricoult SJ, Guarente L. SirT3 suppresses hypoxia inducible factor 1α and tumor growth by inhibiting mitochondrial ROS production. Oncogene. 2011;30(26):2986-96. https://doi.org/10.1038/onc.2011.37 PMid:21358671 PMCid:PMC3134877 |
|
|
89. Semenza GL. Targeting HIF-1 for cancer therapy. Nat Rev Cancer. 2003;3(10):721-32. https://doi.org/10.1038/nrc1187 PMid:13130303 |
|
|
90. Bruick RK, McKnight SL. A conserved family of prolyl-4-hydroxylases that modify HIF. Science. 2001;294(5545):1337-40. https://doi.org/10.1126/science.1066373 PMid:11598268 |
|
|
91. Finley LW, Carracedo A, Lee J, Souza A, Egia A, Zhang J, et al. SIRT3 opposes reprogramming of cancer cell metabolism through HIF1α destabilization. Cancer Cell. 2011;19(3):416-28. https://doi.org/10.1016/j.ccr.2011.02.014 PMid:21397863 PMCid:PMC3065720 |
|
|
92. Quan Y, Wang N, Chen Q, Xu J, Cheng W, Di M, et al. SIRT3 inhibits prostate cancer by destabilizing oncoprotein c-MYC through regulation of the PI3K/Akt pathway. Oncotarget. 2015;6(28):26494-507. https://doi.org/10.18632/oncotarget.4764 PMid:26317998 PMCid:PMC4694917 |
|
|
93. Inuzuka H, Gao D, Finley LW, Yang W, Wan L, Fukushima H, et al. Acetylation-dependent regulation of Skp2 function. Cell. 2012;150(1):179-93. |
|
|
94. Zhang YK, Qu YY, Lin Y, Wu XH, Chen HZ, Wang X, et al. Enoyl-CoA hydratase-1 regulates mTOR signaling and apoptosis by sensing nutrients. Nat Commun. 2017;8(1):464. https://doi.org/10.1038/s41467-017-00489-5 PMid:28878358 PMCid:PMC5587591 |
|
|
95. Lee JJ, van de Ven RAH, Zaganjor E, Ng MR, Barakat A, Demmers JJPG, et al. Inhibition of epithelial cell migration and Src/FAK signaling by SIRT3. Proc Natl Acad Sci U S A. 2018;115(27):7057-62. https://doi.org/10.1073/pnas.1800440115 PMid:29915029 PMCid:PMC6142214 |
|
|
96. Dong XC, Jing LM, Wang WX, Gao YX. Down-regulation of SIRT3 promotes ovarian carcinoma metastasis. Biochem Biophys Res Commun. 2016;475(3):245-50. https://doi.org/10.1016/j.bbrc.2016.05.098 PMid:27216459 |
|
|
97. Li R, Quan Y, Xia W. SIRT3 inhibits prostate cancer metastasis through regulation of FOXO3A by suppressing Wnt/β-catenin pathway. Exp Cell Res. 2018;364(2):143-51. https://doi.org/10.1016/j.yexcr.2018.01.036 PMid:29421536 |
|
|
98. Zeng X, Wang N, Zhai H, Wang R, Wu J, Pu W. SIRT3 functions as a tumor suppressor in hepatocellular carcinoma. Tumour Biol. 2017;39(3):1010428317691178. https://doi.org/10.1177/1010428317691178 PMid:28347248 |
|
|
99. Huang S, Guo H, Cao Y, Xiong J. MiR-708-5p inhibits the progression of pancreatic ductal adenocarcinoma by targeting Sirt3. Pathol Res Pract. 2019;215(4):794-800. https://doi.org/10.1016/j.prp.2019.01.026 PMid:30683474 |
|
|
100. Liu Y, Liu YL, Cheng W, Yin XM, Jiang B. The expression of SIRT3 in primary hepatocellular carcinoma and the mechanism of its tumor suppressing effects. Eur Rev Med Pharmacol Sci. 2017;21(5):978-98. |
|
|
101. Song CL, Tang H, Ran LK, Ko BC, Zhang ZZ, Chen X, et al. Sirtuin 3 inhibits hepatocellular carcinoma growth through the glycogen synthase kinase-3β/BCL2-associated X protein-dependent apoptotic pathway. Oncogene. 2016;35(5):631-41. https://doi.org/10.1038/onc.2015.121 PMid:25915842 |
|
|
102. Allison SJ, Milner J. SIRT3 is pro-apoptotic and participates in distinct basal apoptotic pathways. Cell Cycle. 2007;6(21):2669-77. https://doi.org/10.4161/cc.6.21.4866 PMid:17957139 |
|
|
103. Xiao K, Jiang J, Wang W, Cao S, Zhu L, Zeng H, et al. Sirt3 is a tumor suppressor in lung adenocarcinoma cells. Oncol Rep. 2013;30(3):1323-8. |
|
|
104. Tang X, Li Y, Liu L, Guo R, Zhang P, Zhang Y, et al. Sirtuin 3 induces apoptosis and necroptosis by regulating mutant p53 expression in small‑cell lung cancer. Oncol Rep. 2020;43(2):591-600. https://doi.org/10.3892/or.2019.7439 |
|
|
105. Wu Y, Gao WN, Xue YN, Zhang LC, Zhang JJ, Lu SY, et al. SIRT3 aggravates metformin-induced energy stress and apoptosis in ovarian cancer cells. Exp Cell Res. 2018;367(2):137-49. https://doi.org/10.1016/j.yexcr.2018.03.030 PMid:29580688 |
|
|
106. Xiong Y, Wang M, Zhao J, Wang L, Li X, Zhang Z, et al. SIRT3 is correlated with the malignancy of non-small cell lung cancer. Int J Oncol. 2017;50(3):903-10. https://doi.org/10.3892/ijo.2017.3868 PMid:28197634 |
|
|
107. Cui Y, Qin L, Wu J, Qu X, Hou C, Sun W, et al. SIRT3 Enhances Glycolysis and Proliferation in SIRT3-Expressing Gastric Cancer Cells. PLoS One. 2015;10(6):e0129834. https://doi.org/10.1371/journal.pone.0129834 PMid:26121691 PMCid:PMC4487898 |
|
|
108. Baccelli I, Gareau Y, Lehnertz B, Gingras S, Spinella JF, Corneau S, et al. Mubritinib Targets the Electron Transport Chain Complex I and Reveals the Landscape of OXPHOS Dependency in Acute Myeloid Leukemia. Cancer Cell. 2019;36(1):84-99.e8. https://doi.org/10.1016/j.ccell.2019.06.003 PMid:31287994 |
|
|
109. Li M, Chiang YL, Lyssiotis CA, Teater MR, Hong JY, Shen H, et al. Non-oncogene Addiction to SIRT3 Plays a Critical Role in Lymphomagenesis. Cancer Cell. 2019;35(6):916-31.e9. https://doi.org/10.1016/j.ccell.2019.05.002 PMid:31185214 PMCid:PMC7534582 |
|
|
110. Bergaggio E, Riganti C, Garaffo G, Vitale N, Mereu E, Bandini C, et al. IDH2 inhibition enhances proteasome inhibitor responsiveness in hematological malignancies. Blood. 2019;133(2):156-67. https://doi.org/10.1182/blood-2018-05-850826 PMid:30455381 |
|
|
111. Li H, Feng Z, Wu W, Li J, Zhang J, Xia T. SIRT3 regulates cell proliferation and apoptosis related to energy metabolism in non-small cell lung cancer cells through deacetylation of NMNAT2. Int J Oncol. 2013;43(5):1420-30. https://doi.org/10.3892/ijo.2013.2103 PMid:24042441 PMCid:PMC3823398 |
|
|
112. Chen S, Yang X, Yu M, Wang Z, Liu B, Liu M, et al. SIRT3 regulates cancer cell proliferation through deacetylation of PYCR1 in proline metabolism. Neoplasia. 2019;21(7):665-75. https://doi.org/10.1016/j.neo.2019.04.008 PMid:31108370 PMCid:PMC6526305 |
|
|
113. Wei Z, Song J, Wang G, Cui X, Zheng J, Tang Y, et al. Deacetylation of serine hydroxymethyl-transferase 2 by SIRT3 promotes colorectal carcinogenesis. Nat Commun. 2018;9(1):4468. https://doi.org/10.1038/s41467-018-06812-y PMid:30367038 PMCid:PMC6203763 |
|
|
114. Wang Y, Sun X, Ji K, Du L, Xu C, He N, et al. Sirt3-mediated mitochondrial fission regulates the colorectal cancer stress response by modulating the Akt/PTEN signalling pathway. Biomed Pharmacother. 2018;105:1172-82. https://doi.org/10.1016/j.biopha.2018.06.071 PMid:30021354 |
|
|
115. Xu LX, Hao LJ, Ma JQ, Liu JK, Hasim A. SIRT3 promotes the invasion and metastasis of cervical cancer cells by regulating fatty acid synthase. Mol Cell Biochem. 2020;464(1-2):11-20. https://doi.org/10.1007/s11010-019-03644-2 PMid:31677030 |
|
|
116. Kamarajan P, Alhazzazi TY, Danciu T, D'silva NJ, Verdin E, Kapila YL. Receptor-interacting protein (RIP) and Sirtuin-3 (SIRT3) are on opposite sides of anoikis and tumorigenesis. Cancer. 2012;118(23):5800-10. https://doi.org/10.1002/cncr.27655 PMid:22674009 PMCid:PMC3443499 |
|
|
117. Sengupta A, Haldar D. Human sirtuin 3 (SIRT3) deacetylates histone H3 lysine 56 to promote nonhomologous end joining repair. DNA Repair (Amst). 2018;61:1-16. https://doi.org/10.1016/j.dnarep.2018.04.001 https://doi.org/10.1016/j.dnarep.2017.11.003 PMid:29136592 |
|
|
118. Carafa V, Rotili D, Forgione M, Cuomo F, Serretiello E, Hailu GS, et al. Sirtuin functions and modulation: from chemistry to the clinic. Clin Epigenetics. 2016;8:61. https://doi.org/10.1186/s13148-016-0224-3 PMid:27226812 PMCid:PMC4879741 |
|
|
119. Zhang J, Xiang H, Liu J, Chen Y, He RR, Liu B. Mitochondrial Sirtuin 3: New emerging biological function and therapeutic target. Theranostics. 2020;10(18):8315-42. https://doi.org/10.7150/thno.45922 PMid:32724473 PMCid:PMC7381741 |
|
|
120. Rajman L, Chwalek K, Sinclair DA. Therapeutic Potential of NAD-Boosting Molecules: The In Vivo Evidence. Cell Metab. 2018;27(3):529-47. |
|
|
121. Santidrian AF, Matsuno-Yagi A, Ritland M, Seo BB, LeBoeuf SE, Gay LJ, et al. Mitochondrial complex I activity and NAD+/NADH balance regulate breast cancer progression. J Clin Invest. 2013;123(3):1068-81. https://doi.org/10.1172/JCI64264 PMid:23426180 PMCid:PMC3582128 |
|
|
122. Pillai VB, Samant S, Sundaresan NR, Raghuraman H, Kim G, Bonner MY, et al. Honokiol blocks and reverses cardiac hypertrophy in mice by activating mitochondrial Sirt3. Nat Commun. 2015;6:6656. https://doi.org/10.1038/ncomms7656 PMid:25871545 PMCid:PMC4441304 |
|
|
123. Arora S, Singh S, Piazza GA, Contreras CM, Panyam J, Singh AP. Honokiol: a novel natural agent for cancer prevention and therapy. Curr Mol Med. 2012;12(10):1244-52. https://doi.org/10.2174/156652412803833508 PMid:22834827 PMCid:PMC3663139 |
|
|
124. Xu HL, Tang W, Du GH, Kokudo N. Targeting apoptosis pathways in cancer with magnolol and honokiol, bioactive constituents of the bark of Magnolia officinalis. Drug Discov Ther. 2011;5(5):202-10. https://doi.org/10.5582/ddt.2011.v5.5.202 PMid:22466367 |
|
|
125. Arora S, Bhardwaj A, Srivastava SK, Singh S, McClellan S, Wang B, et al. Honokiol arrests cell cycle, induces apoptosis, and potentiates the cytotoxic effect of gemcitabine in human pancreatic cancer cells. PLoS One. 2011;6(6):e21573. https://doi.org/10.1371/journal.pone.0021573 PMid:21720559 PMCid:PMC3123370 |
|
|
126. Pillai VB, Kanwal A, Fang YH, Sharp WW, Samant S, Arbiser J, et al. Honokiol, an activator of Sirtuin-3 (SIRT3) preserves mitochondria and protects the heart from doxorubicin-induced cardiomyopathy in mice. Oncotarget. 2017;8(21):34082-98. https://doi.org/10.18632/oncotarget.16133 PMid:28423723 PMCid:PMC5470953 |
|
|
127. Tian W, Deng Y, Li L, He H, Sun J, Xu D. Honokiol synergizes chemotherapy drugs in multidrug resistant breast cancer cells via enhanced apoptosis and additional programmed necrotic death. Int J Oncol. 2013;42(2):721-32. https://doi.org/10.3892/ijo.2012.1739 PMid:23242346 |
|
|
128. Huang K, Chen Y, Zhang R, Wu Y, Ma Y, Fang X, et al. Honokiol induces apoptosis and autophagy via the ROS/ERK1/2 signaling pathway in human osteosarcoma cells in vitro and in vivo. Cell Death Dis. 2018;9(2):157. https://doi.org/10.1038/s41419-017-0166-5 PMid:29410403 PMCid:PMC5833587 |
|
|
129. Li Y, Ye Z, Lai W, Rao J, Huang W, Zhang X, et al. Activation of Sirtuin 3 by Silybin Attenuates Mitochondrial Dysfunction in Cisplatin-induced Acute Kidney Injury. Front Pharmacol. 2017;8:178. https://doi.org/10.3389/fphar.2017.00178 PMid:28424621 PMCid:PMC5380914 |
|
|
130. Loguercio C, Festi D. Silybin and the liver: from basic research to clinical practice. World J Gastroenterol. 2011;17(18):2288-301. https://doi.org/10.3748/wjg.v17.i18.2288 PMid:21633595 PMCid:PMC3098397 |
|
|
131. Kaur M, Velmurugan B, Tyagi A, Deep G, Katiyar S, Agarwal C, et al. Silibinin suppresses growth and induces apoptotic death of human colorectal carcinoma LoVo cells in culture and tumor xenograft. Mol Cancer Ther. 2009;8(8):2366-74. https://doi.org/10.1158/1535-7163.MCT-09-0304 PMid:19638451 PMCid:PMC2728169 |
|
|
132. Flaig TW, Glodé M, Gustafson D, van Bokhoven A, Tao Y, Wilson S, et al. A study of high-dose oral silybin-phytosome followed by prostatectomy in patients with localized prostate cancer. Prostate. 2010;70(8):848-55. https://doi.org/10.1002/pros.21118 PMid:20127732 |
|
|
133. Gertz M, Nguyen GT, Fischer F, Suenkel B, Schlicker C, Fränzel B, et al. A molecular mechanism for direct sirtuin activation by resveratrol. PLoS One. 2012;7(11):e49761. https://doi.org/10.1371/journal.pone.0049761 PMid:23185430 PMCid:PMC3504108 |
|
|
134. Pan W, Yu H, Huang S, Zhu P. Resveratrol Protects against TNF-α-Induced Injury in Human Umbilical Endothelial Cells through Promoting Sirtuin-1-Induced Repression of NF-KB and p38 MAPK. PLoS One. 2016;11(1):e0147034. https://doi.org/10.1371/journal.pone.0147034 PMid:26799794 PMCid:PMC4723256 |
|
|
135. Howitz KT, Bitterman KJ, Cohen HY, Lamming DW, Lavu S, Wood JG, et al. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature. 2003;425(6954):191-6. https://doi.org/10.1038/nature01960 PMid:12939617 |
|
|
136. Kershaw J, Kim KH. The Therapeutic Potential of Piceatannol, a Natural Stilbene, in Metabolic Diseases: A Review. J Med Food. 2017;20(5):427-38. |
|
|
137. Bagul PK, Katare PB, Bugga P, Dinda AK, Banerjee SK. SIRT-3 Modulation by Resveratrol Improves Mitochondrial Oxidative Phosphorylation in Diabetic Heart through Deacetylation of TFAM. Cells. 2018;7(12). https://doi.org/10.3390/cells7120235 PMid:30487434 PMCid:PMC6315986 |
|
|
138. Zhou X, Chen M, Zeng X, Yang J, Deng H, Yi L, et al. Resveratrol regulates mitochondrial reactive oxygen species homeostasis through Sirt3 signaling pathway in human vascular endothelial cells. Cell Death Dis. 2014;5:e1576. https://doi.org/10.1038/cddis.2014.530 PMid:25522270 PMCid:PMC4454164 |
|
|
139. Guan X, Lin P, Knoll E, Chakrabarti R. Mechanism of inhibition of the human sirtuin enzyme SIRT3 by nicotinamide: computational and experimental studies. PLoS One. 2014;9(9):e107729. https://doi.org/10.1371/journal.pone.0107729 PMid:25221980 PMCid:PMC4164625 |
|
|
140. Nikas IP, Paschou SA, Ryu HS. The Role of Nicotinamide in Cancer Chemoprevention and Therapy. Biomolecules. 2020;10(3). https://doi.org/10.3390/biom10030477 PMid:32245130 PMCid:PMC7175378 |
|
|
141. Galli U, Mesenzani O, Coppo C, Sorba G, Canonico PL, Tron GC, et al. Identification of a sirtuin 3 inhibitor that displays selectivity over sirtuin 1 and 2. Eur J Med Chem. 2012;55:58-66. https://doi.org/10.1016/j.ejmech.2012.07.001 PMid:22835719 |
|
|
142. Chen G, Zhang B, Xu H, Sun Y, Shi Y, Luo Y, et al. Suppression of Sirt1 sensitizes lung cancer cells to WEE1 inhibitor MK-1775-induced DNA damage and apoptosis. Oncogene. 2017;36(50):6863-72. https://doi.org/10.1038/onc.2017.297 PMid:28869605 |
|
|
143. Broussy S, Laaroussi H, Vidal M. Biochemical mechanism and biological effects of the inhibition of silent information regulator 1 (SIRT1) by EX-527 (SEN0014196 or selisistat). J Enzyme Inhib Med Chem. 2020;35(1):1124-36.https://doi.org/10.1080/14756366.2020.1758691 PMid:32366137 PMCid:PMC7241506 |
|
|
144. Süssmuth SD, Haider S, Landwehrmeyer GB, Farmer R, Frost C, Tripepi G, et al. An exploratory double-blind, randomized clinical trial with selisistat, a SirT1 inhibitor, in patients with Huntington's disease. Br J Clin Pharmacol. 2015;79(3):465-76. https://doi.org/10.1111/bcp.12512 PMid:25223731 PMCid:PMC4345957 |
|
|
145. Nguyen GT, Gertz M, Steegborn C. Crystal structures of Sirt3 complexes with 4'-bromo-resveratrol reveal binding sites and inhibition mechanism. Chem Biol. 2013;20(11):1375-85. https://doi.org/10.1016/j.chembiol.2013.09.019 PMid:24211137 |
|
|
146. George J, Nihal M, Singh CK, Ahmad N. 4'-Bromo-resveratrol, a dual Sirtuin-1 and Sirtuin-3 inhibitor, inhibits melanoma cell growth through mitochondrial metabolic reprogramming. Mol Carcinog. 2019;58(10):1876-85. https://doi.org/10.1002/mc.23080 PMid:31292999 PMCid:PMC6721992 |
|
|
147. Karaman B, Sippl W. Docking and binding free energy calculations of sirtuin inhibitors. Eur J Med Chem. 2015;93:584-98. https://doi.org/10.1016/j.ejmech.2015.02.045 PMid:25748123 |
|
|
148. Patel K, Sherrill J, Mrksich M, Scholle MD. Discovery of SIRT3 Inhibitors Using SAMDI Mass Spectrometry. J Biomol Screen. 2015;20(7):842-8. |
|
|
149. Lain S, Hollick JJ, Campbell J, Staples OD, Higgins M, Aoubala M, et al. Discovery, in vivo activity, and mechanism of action of a small-molecule p53 activator. Cancer Cell. 2008;13(5):454-63. https://doi.org/10.1016/j.ccr.2008.03.004 PMid:18455128 PMCid:PMC2742717 |
|
|
150. Hirai S, Endo S, Saito R, Hirose M, Ueno T, Suzuki H, et al. Antitumor effects of a sirtuin inhibitor, tenovin-6, against gastric cancer cells via death receptor 5 up-regulation. PLoS One. 2014;9(7):e102831. https://doi.org/10.1371/journal.pone.0102831 PMid:25033286 PMCid:PMC4102575 |
|
|
151. Rousalova I, Banerjee S, Sangwan V, Evenson K, McCauley JA, Kratzke R, et al. Minnelide: a novel therapeutic that promotes apoptosis in non-small cell lung carcinoma in vivo. PLoS One. 2013;8(10):e77411. https://doi.org/10.1371/journal.pone.0077411 PMid:24143232 PMCid:PMC3797124 |
|
|
152. Kumar A, Corey C, Scott I, Shiva S, D'Cunha J. Minnelide/Triptolide Impairs Mitochondrial Function by Regulating SIRT3 in P53-Dependent Manner in Non-Small Cell Lung Cancer. PLoS One. 2016;11(8):e0160783. https://doi.org/10.1371/journal.pone.0160783 PMid:27501149 PMCid:PMC4976872 |
|
|
153. Gorska-Ponikowska M, Kuban-Jankowska A, Eisler SA, Perricone U, Lo Bosco G, Barone G, et al. 2-Methoxyestradiol Affects Mitochondrial Biogenesis Pathway and Succinate Dehydrogenase Complex Flavoprotein Subunit A in Osteosarcoma Cancer Cells. Cancer Genomics Proteomics. 2018;15(1):73-89. |
|
|
154. Disch JS, Evindar G, Chiu CH, Blum CA, Dai H, Jin L, et al. Discovery of thieno[3,2-d]pyrimidine-6-carboxamides as potent inhibitors of SIRT1, SIRT2, and SIRT3. J Med Chem. 2013;56(9):3666-79. https://doi.org/10.1021/jm400204k PMid:23570514 |
|
|
155. Zhou Y, Li C, Peng J, Xie L, Meng L, Li Q, et al. DNA-Encoded Dynamic Chemical Library and Its Applications in Ligand Discovery. J Am Chem Soc. 2018;140(46):15859-67. https://doi.org/10.1021/jacs.8b09277 PMid:30412395 |
|
|
156. Ma J, Liu B, Yu D, Zuo Y, Cai R, Yang J, et al. SIRT3 deacetylase activity confers chemoresistance in AML via regulation of mitochondrial oxidative phosphorylation. Br J Haematol. 2019;187(1):49-64. https://doi.org/10.1111/bjh.16044 PMid:31236919 PMCid:PMC6790595 |
|
|
157. Schaefer PM, Hilpert D, Niederschweiberer M, Neuhauser L, Kalinina S, Calzia E, et al. Mitochondrial matrix pH as a decisive factor in neurometabolic imaging. Neurophotonics. 2017;4(4):045004. https://doi.org/10.1117/1.NPh.4.4.045004 PMid:29181426 PMCid:PMC5685807 |
|
|
158. Schaefer PM, Kalinina S, Rueck A, von Arnim CAF, von Einem B. NADH Autofluorescence-A Marker on its Way to Boost Bioenergetic Research. Cytometry A. 2018 https://doi.org/10.1002/cyto.a.23597 PMid:30211978 |